| |
Extrahepatic replication of HCV: Insights into clinical manifestations and biological consequences
|
| |
| |
Hepatology July 2006
Volume 44, Issue 1, Pages 15-22
Jason T. Blackard *, Nyingi Kemmer, Kenneth E. Sherman
Division of Digestive Diseases, University of Cincinnati College of Medicine, Cincinnati, OH
Abstract
An estimated 170 million persons are infected with the hepatitis C virus (HCV) worldwide. While hepatocytes are the major site of infection, a broad clinical spectrum of extrahepatic complications and diseases are associated with chronic HCV infection, highlighting the involvement of HCV in a variety of non-hepatic pathogenic processes. There is a growing body of evidence to suggest that HCV can replicate efficiently in extrahepatic tissues and cell types, including peripheral blood mononuclear cells. Nonetheless, laboratory confirmation of HCV replication in extrahepatic sites is fraught with technical challenges, and in vitro systems to investigate extrahepatic replication of HCV are severely limited. Thus, future studies of extrahepatic replication should combine innovative in vitro assays with a prospective cohort design to maximize our understanding of this important phenomenon to the pathogenesis and treatment response rates of HCV.
Article Text
An estimated 170 million persons are infected with the hepatitis C virus (HCV) worldwide. Infection with HCV is a major cause of chronic liver disease, with >75% of patients developing chronic hepatitis and 20% of chronically infected persons developing cirrhosis.[1][2] HCV infection is also a major cause of hepatocellular carcinoma and the primary reason for liver transplantation in the United States. While hepatocytes are the major site of viral replication, a broad clinical spectrum of extrahepatic complications and diseases are associated with chronic HCV infection,[3-6] including mixed cryoglobulinemia, non-Hodgkin's lymphoma, cutaneous vasculitis, glomerulonephritis, neuropathy, and lymphoproliferative disorders to highlight just a few (Table 1). Extrahepatic manifestations are quite common among patients infected with HCV. In a large prospective cohort study, the MULTIVIRC group reported that 74% of patients had a least one clinical extrahepatic manifestation.[7] The most commonly identified manifestations were mixed cryoglobulinemia (MC) (40%), arthralgia (23%), paresthesia (17%), myalgia (15%), pruritus (15%), and sicca syndrome (11%). Several of the most clinically significant extrahepatic manifestations of HCV are as follows:
Table 1. Clinical Manifestations Associated With HCV Disease
Mixed cryoglobulinemia
Non-Hodgkin's lymphoma
Cutaneous vasculitis
Glomerulonephritis
Neuropathy
Lymphoproliferative disorders
Porphyria cutanea tarda
Lichen planus
Sjogren's syndrome
Detection of Extrahepatic Replication
Because positive-sense HCV RNA resides within individual virions, the mere detection of positive-sense HCV RNA in cell preparations does not definitively prove replication. Thus, the most important indicator of HCV genomic replication is the production of negative-strand HCV RNA, as sustained viral replication within a given cellular environment can only be maintained through the constant production of replicative intermediate molecules.[33] A predominance of positive-strand over negative-strand HCV RNA occurs during intrahepatic replication of HCV[34-36]; however, the various factors that control this asymmetry are not well described, and this phenomenon has not been adequately characterized in non-hepatic cell types.
While traditional HCV viral load assays quantify HCV RNA in the serum, they are unable to distinguish positive- from negative-strand HCV RNA and, thus have limited utility in assessing extrahepatic replication. Nonetheless, highly sensitive detection methods are continuously being developed and should facilitate a more accurate assessment of the true extent of HCV infection/replication in a variety of cell types and tissues.[37][38] To date, extrahepatic replication of HCV has been explored using a plethora of cell culture and molecular biological assays. The use of strand-specific reverse transcriptase polymerase chain reaction (RT-PCR) to detect negative-strand HCV RNA is often cited as evidence for viral replication. However, due to the potential for contamination and the lack of strand-specificity, standard RT-PCR assays should not be used to characterize extrahepatic replication of HCV.[33] A new approach has been to enhance the strand specificity of RT-PCR through the use of recombinant Tth enzyme, which has independent reverse transcriptase and polymerase activities.[39] Because of the 10,000-fold ability of Tth to discriminate between specific strands, it is ideal for the specific detection of negative-strand HCV RNA and has quickly replaced several of the previous, less-specific PCR-based strategies. Real-time, strand-specific PCR assays utilizing Tth are also now available.[35][40][41]
Recently, the development of self-replicating HCV RNA replicons has revolutionized the study of HCV;[42] however, these systems are restricted to a single, highly permissive hepatocyte-derived cell line.[43][44] Only within the last year have HCV systems that support infectious HCV production been reported,[45-48] although they do not support HCV replication in non-hepatocyte-derived cells. Consequently, additional systems to study sustained viral replication in multiple hepatocyte- and non-hepatocyte-derived cells are critically needed to advance the molecular characterization of extrahepatic replication of HCV and its role in the progression of the disease.
Several complementary strategies to investigate extrahepatic replication of HCV include DNA-based expression systems,[41][49-53] lymphocyte cell lines that constitutively express HCV,[21][23][54] peripheral blood mononuclear cell (PBMC) cultures from HCV-infected persons,[21][55][56] direct viral infection of extrahepatic cell types with HCV-infected sera,[35][57-60] in situ hybridization and immunohistochemistry,[38][61-64] and laser capture microdissection.[37][65] The relevance of each of these strategies to extrahepatic replication of HCV is not currently known and can only be determined through extensive validation and cross-comparison amongst all available methodologies.
Evidence for extrahepatic Replication of HCV
HCV binds several cell surface receptors,[66] although cell tropism and the specific host determinants required for HCV genome replication are not well characterized. Thus, it is difficult to determine a priori which cell types will support HCV replication, as indicated by the synthesis and accumulation of HCV negative-strand RNAs. Despite these limitations, extrahepatic replication of HCV has been documented extensively in vivo. For instance, negative-strand HCV RNA has been identified in a variety of non-hepatic tissues (Table 2), including the pancreas, thyroid, bone marrow, adrenal gland, spleen, lymph node, cervicovaginal fluid, and brain.[29][67-72]
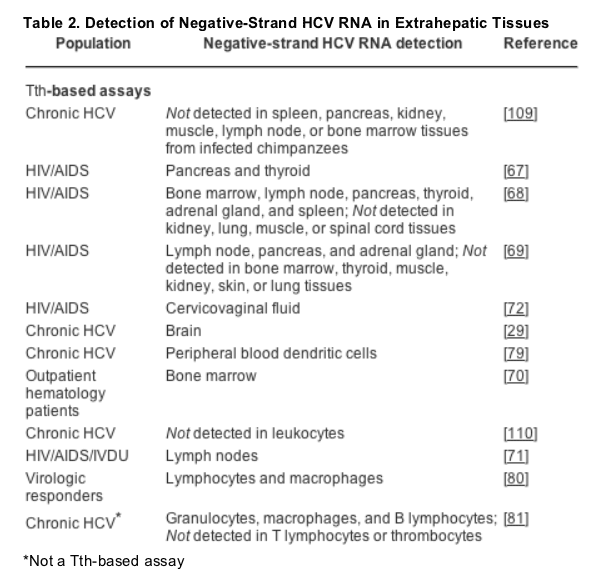
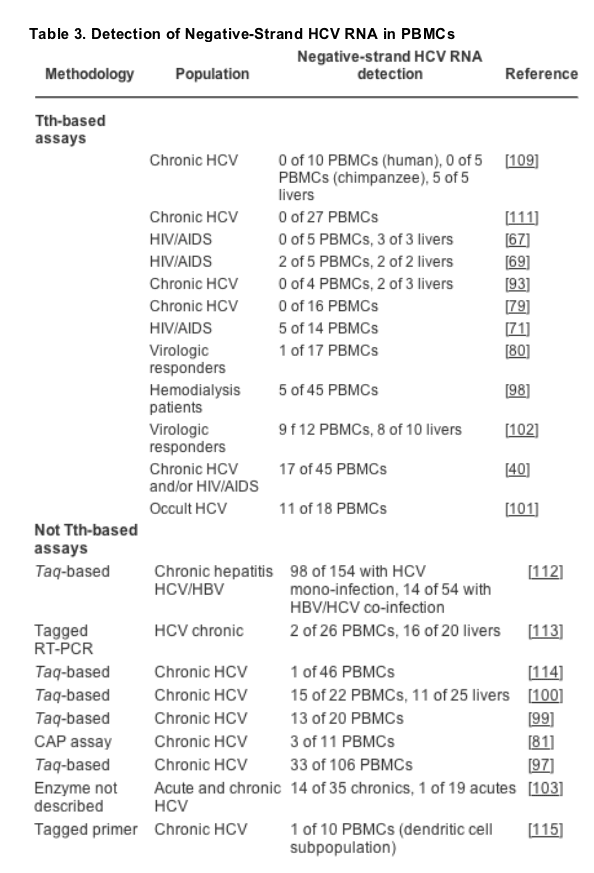
Initial studies similarly reported extrahepatic replication of HCV in PBMCs (Table 3).[58][73-77] However, as the detection assays were not able to adequately differentiate between positive- and negative-strand HCV RNAs, theses results should be interpreted with extreme caution. Subsequent studies have now convincingly demonstrated negative-strand HCV RNA - albeit in a minority of PBMCs tested - particularly among immunosuppressed patients, such as HIV/AIDS patients[68][69][71] and liver transplant recipients.[63][78] There is also a growing body of evidence to suggest that HCV replication may occur within peripheral dendritic cells, granulocytes, B lymphocytes, and monocytes/macrophages.[79-81] While some of these data should be viewed cautiously, as a variety of techniques were utilized and a limited number of samples were analyzed, they are intriguing in light of the extrahepatic manifestations of HCV reviewed earlier.
Pioneering studies by Shimizu et al. demonstrated that HCV infection of lymphocytes occurs both in vitro and in vivo.[82-84] Interestingly, sequence analysis has subsequently revealed that certain viral variants may be selected for growth in extrahepatic cell types, implying that HCV diversity directly impacts cell tropism.[85][86] Furthermore, several studies have described a non-random distribution of HCV sequences in hepatic and extrahepatic compartments,[72][87-96] leading to the conclusion that the presence of tissue-specific sequences is compatible with independent viral replication in extrahepatic sites. It has also been speculated that HCV variants within distinct compartments may differ in their sensitivity to interferon, although this hypothesis has not been formally tested. Viral sequence evolution after HCV therapy cessation could also suggest emergence from a protected compartment, although this would require extensive characterization of viruses before and after treatment in multiple tissue/cellular compartments. The potential correlation between extrahepatic replication and HCV genotype has also been examined,[81][97] with at least one study suggesting that negative-strand HCV RNA may be more frequently detected in the PBMCs from HCV genotype 1-infected persons.[81] Nonetheless, these data will require rigorous validation in larger clinical populations and additional follow-up data to determine if certain HCV genotypes are more frequently associated with the development of specific extrahepatic manifestations.
Clinical Implications of Extrahepatic Replication
To date, prospective studies of extrahepatic replication are missing from the available literature. Nonetheless, retrospective studies suggest several potential associations between extrahepatic replication of HCV and important clinical outcomes. For example, several studies have found that HCV RNA may be present in PBMCs but not the corresponding serum or liver.[98-101] Thus, current detection systems may underestimate the true extent of replication. Moreover, HCV RNA, including replicative intermediate forms, can persist at very low levels in peripheral lymphoid cells for many years after apparently complete spontaneous or treatment-induced resolution of chronic HCV,[102] strongly suggesting that sub-populations within PBMCs may represent true reservoirs of HCV replication. Interestingly, patients with detectable negative-strand HCV RNA in PBMCs had lower interferon (IFN) sustained response rates compared to those without detectable negative-strand HCV RNA in PBMCs.[103] Thus, it is provocative to speculate that low-level replication of HCV in PBMCs may lead to reactivation of HCV after termination of therapy and/or predict response to therapy.[80][99][100][102-105]
As reviewed elsewhere, HCV is likely involved in several neurologic syndromes.[16] While central nervous system (CNS) involvement is less frequent, the detection of negative-strand HCV RNA in the CNS[29] suggests a potential link between HCV and these extrahepatic pathologies. However, the underlying biological mechanisms have not been elucidated. Similarly, the detection of HCV RNA in cervical lymphocytes[106] and HCV compartmentalization within cervicovaginal fluid[72] suggests that this compartment may contribute to the sexual transmission of HCV. Nonetheless, few studies have examined the temporal relationship between HCV RNA levels in extrahepatic compartments and the development of specific extrahepatic complications, although successful treatment of HCV is frequently associated with resolution of several extrahepatic complications of HCV infection.[8][9][13][16][26]
The proposed pathogenic processes by which HCV may result in the extrahepatic manifestations described above have been described in detail elsewhere.[4-6][107] Importantly, multiple immunologic, virologic, and environmental factors are likely involved. For instance, given that many extrahepatic manifestations are lymphoproliferative disorders and/or autoimmune disorders, it is tempting to speculate that the lymphotropism of HCV may be an important contributing factor. Moreover, it has long been hypothesized that chronic antigen stimulation by HCV activates B cells and may ultimately result in cryoglobulinemia and non-Hodgkin lymphoma.[5] Furthermore, specific genomic regions, such as the 5UTR and HVR-1, have also been identified that may impact cell tropism,[85][86] thereby influencing the extent of extrahepatic replication. There is also evidence to suggest that the HCV core protein can regulate various cellular oncogenes and viral promoters that regulate transformation and/or carcinogenesis.[108]
Extrahepatic Replication: the Research Road Ahead
Until recently, the existence of extrahepatic reservoirs of HCV has been somewhat controversial due to conflicting findings in vivo. A number of current reports now suggest that extrahepatic replication of HCV does indeed occur and that it may have profound effects on HCV treatment and disease pathogenesis. Nevertheless, a focused research agenda is still lacking with several critical research areas remaining to be explored.
While epidemiological studies provide important data regarding possible associations, it is essential that the link between HCV replication and development of extrahepatic diseases be examined at a mechanistic level to understand the precise pathogenic processes that underlie these clinical outcomes. This is a large field of research that includes characterization of the pathogenic mechanisms by which HCV can lead to the development of lymphomas, the mechanisms that link HCV to insulin resistance and diabetes, and the association of HCV with autoimmune-mediated thyroid diseases. Additional prospective studies could further define the true extent of extrahepatic replication of HCV and the subsequent development of extrahepatic manifestations by measuring multiple virological, immunological, and clinical parameters in large, well-characterized cohorts. In the absence of robust culture systems that permit detailed examinations of HCV replication in extrahepatic cell types, such studies are also absolutely critical to demonstrate a causal relationship between viral replication in specific extrahepatic compartments and the subsequent development of certain extrahepatic manifestations of HCV. Moreover, several groups have suggested that extrahepatic replication is associated with treatment outcome, although this has not been formally tested in large clinical cohorts receiving HCV therapy. In such cohorts, the association of baseline detection of negative-strand HCV RNA, as well as a more rigorous examination of HCV viral variants present in extrahepatic compartments, should be examined as possible predictors of treatment failure. Finally, additional in vitro replication systems that allow direct comparison of HCV replication among a variety of cell types of hepatic and extrahepatic origin will be absolutely essential to elucidate the complex pathogenic processes by which extrahepatic HCV occurs and results in a myriad of disease manifestations.
Cutaneous Vasculitis
Intermittent purpura of the lower extremities is the most common clinical finding and is a direct consequence of immune complex deposition in the dermal capillaries. A sensation of cutaneous burns occasionally precedes its appearance. It is usually benign and self-limited with resolution of symptoms within a week, although the vasculitic process will progress to necrotizing skin lesions with ulcerations in approximately 10% of patients. The severity of vasculitis correlates with the level of HCV viremia but not with the level of serum cryoglobulins. Skin biopsy confirms the diagnosis with the classic finding of leukocytoclastic vasculitis. Cutaneous vasculitis is the MC manifestation that responds best to antiviral therapy with pegylated interferon (PEG-IFN) and ribavirin (RBV).[8][9]
Cutaneous Vasculitis
Intermittent purpura of the lower extremities is the most common clinical finding and is a direct consequence of immune complex deposition in the dermal capillaries. A sensation of cutaneous burns occasionally precedes its appearance. It is usually benign and self-limited with resolution of symptoms within a week, although the vasculitic process will progress to necrotizing skin lesions with ulcerations in approximately 10% of patients. The severity of vasculitis correlates with the level of HCV viremia but not with the level of serum cryoglobulins. Skin biopsy confirms the diagnosis with the classic finding of leukocytoclastic vasculitis. Cutaneous vasculitis is the MC manifestation that responds best to antiviral therapy with pegylated interferon (PEG-IFN) and ribavirin (RBV).[8][9]
Glomerulonephritis
The association between HCV infection and renal disease is well established. Membranoproliferative glomerulonephritis (MPGN) is the most common extrahepatic manifestation of HCV that involves the kidneys, representing 70 to 80% of HCV-associated glomerulonephritis. The prevalence of HCV infection in MPGN is 10 to 20% in the United States.[10] There are two forms of HCV-associated MPGN described in the literature: cryoglobulin-related and non-cryoglobulin renal disease. Irrespective of the form, the most common presentation is proteinuria and microscopic hematuria.[11] The clinical course of MPGN is variable with approximately 30% of patients having complete or partial remission, 30% with intermittent exacerbation and remission, 30% with an indolent course, and 10% progressing to chronic renal failure.[12] Therapy with PEG-IFN + RBV has proven to be effective in some patients with a significant improvement in proteinuria and serum creatinine levels.[13] Other therapies have achieved variable clinical response rates.[14][15]
Neuropathy
Neurologic manifestations in HCV-infected patients occur predominantly in the peripheral nervous system and may affect as many as 50% of cases.[16] The prevalence of peripheral nerve involvement in patients with MC ranges between 20 and 35%, with clinical manifestations resulting from immune complex deposits within vasa nervorum of the peripheral nerves leading to vasculitis.[17][18] At least 80% with neurologic manifestations present with peripheral neuropathy in the form of a symmetrical distal polyneuropathy that involves the legs and is typically very painful, with progression to muscle weakness in some patients. The clinical course is progressive and response to antiviral therapy and plasmapheresis has been discouraging.[19]
Lymphoproliferative Disorders
Recent studies have found a high prevalence of lymphoproliferative disorders (LPD) in HCV-infected patients.[20] Although there is limited data to suggest that HCV replication occurs in B lymphocytes,[21-23] chronic antigenic stimulation by the virus may trigger B cell proliferation resulting in a wide spectrum of pathology ranging from minor expansion of B cell populations to an aggressive high-grade lymphoma. A high prevalence of HCV in patients with B cell non-Hodgkin's lymphoma (NHL) has been reported in some, but not all studies.[24] A recent meta-analysis has also confirmed this strong positive association between HCV and the risk of NHL,[20] although mechanistic studies are still lacking. In HCV-infected patients, the vast majority of NHL is low grade with predominantly extranodal involvement of organs such as the liver, spleen, salivary glands, and stomach.[25] The optimal treatment of the HCV-related B-cell NHL is not clear; however, emerging data of remission of LPD after HCV eradication in some subsets of splenic and gastric mucosa-associated lymphoid tissue (MALT) lymphomas are encouraging.[26]
Over the last decade, epidemiological data have demonstrated a possible link between porphyria cutanea tarda, lichen planus, Sjogren's syndrome and HCV.[27] More recently, HCV has been associated with fatigue[28] and insulin resistance though this association is weak.[29][30] HCV has also been implicated in the pathogenesis of acquired aplastic anemia, although this hematological disorder is uncommon in HCV infected individuals and the available evidence to support a causal relationship is very limited.[31][32] A greater knowledge of HCV replication in non-hepatic organs will undoubtedly provide useful insights into the pathogenic processes by which HCV is involved in these extrahepatic manifestations.
References
1 Seeff L. Natural history of viral hepatitis, type C. Semin Gastrointest Dis 1995; 6: 20-27. Links
2 Lauer G, Walker B. Medical progress: hepatitis C virus infection. N Engl J Med 2001; 345: 41-52. Links
3 Gumber S, Chopra S. Hepatitis C: a multifaceted disease-review of extrahepatic manifestations. Ann Intern Med 1995; 123: 615-620. Links
4 Agnello V, De Rosa FG. Extrahepatic disease manifestations of HCV infection: some current issues. J Hepatol 2004; 40: 341-352. Links
5 Mayo M. Extrahepatic manifestations of hepatitis C infection. Am J Med Sci 2002; 325: 135-148. Links
6 Sene D, Limal N, Cacoub P. Hepatitis C virus-associated extrahepatic manifestations: a review. Metab Brain Dis 2004; 19: 357-381. Links
7 Cacoub P, Poynard T, Ghillani P, Charlotte F, Olivi M, Piette JC, et al. Extrahepatic manifestations of chronic hepatitis C. MULTIVIRC Group. Multidepartment Virus C. Arthritis Rheum 1999; 42: 2204-2212. Links
8 Ferri C, Sebastiani M, Giuggioli D, Cazzato M, Longombardo G, Antonelli A, et al. Mixed cryoglobulinemia: demographic, clinical, and serologic features and survival in 231 patients. Semin Arthritis Rheum 2004; 33: 355-374. Links
9 Cacoub P, Saadoun D, Limal N, Sene D, Lidove O, Piette JC. PEGylated interferon alfa-2b and ribavirin treatment in patients with hepatitis C virus-related systemic vasculitis. Arthritis Rheum 2005; 52: 911-915. Links
10 Johnson RJ, Gretch DR, Yamabe H, Hart J, Bacchi CE, Hartwell P, et al. Membranoproliferative glomerulonephritis associated with hepatitis C virus infection. N Engl J Med 1993; 328: 465-470. Links
11 Daghestani L, Pomeroy C. Renal manifestations of hepatitis C infection. Am J Med 1999; 106: 347-354. Links
12 D'Amico G, Fornasieri A. Cryoglobulinemic glomerulonephritis: a membranoproliferative glomerulonephritis induced by hepatitis C virus. Am J Kidney Dis 1995; 25: 361-369. Links
13 Alric L, Plaisier E, Thebault S, Peron JM, Rostaing L, Pourrat J, et al. Influence of antiviral therapy in hepatitis C virus-associated cryoglobulinemic MPGN. Am J Kidney Dis 2004; 43: 617-623. Links
14 Garini G, Allegri L, Vaglio A, Buzio C. Hepatitis C virus-related cryoglobulinemia and glomerulonephritis: pathogenesis and therapeutic strategies. Ann Ital Med Int 2005; 20: 71-80. Links
15 Meyers CM, Seeff LB, Stehman-Breen CO, Hoofnagle JH. Hepatitis C and renal disease: an update. Am J Kidney Dis 2003; 42: 631-657. Links
16 Cacoub P, Saadoun D, Limal N, Leger JM, Maisonobe T. Hepatitis C virus infection and mixed cryoglobulinaemia vasculitis: a review of neurological complications. AIDS 2005; 19 Suppl 3: S128-134. Links
17 Bonetti B, Scardoni M, Monaco S, Rizzuto N, Scarpa A. Hepatitis C virus infection of peripheral nerves in type II cryoglobulinaemia. Virchows Arch 1999; 434: 533-535. Links
18 De Martino L, Sampaolo S, Tucci C, Ambrosone L, Budillon A, Migliaresi S, et al. Viral RNA in nerve tissues of patients with hepatitis C infection and peripheral neuropathy. Muscle Nerve 2003; 27: 102-104. Links
19 Ramos-Casals M, Trejo O, Garcia-Carrasco M, Font J. Therapeutic management of extrahepatic manifestations in patients with chronic hepatitis C virus infection. Rheumatology (Oxford) 2003; 42: 818-828. Links
20 Matsuo K, Kusano A, Sugumar A, Nakamura S, Tajima K, Mueller NE. Effect of hepatitis C virus infection on the risk of non-Hodgkin's lymphoma: a meta-analysis of epidemiological studies. Cancer Sci 2004; 95: 745-752. Links
21 Bare P, Massud I, Parodi C, Belmonte L, Garcia G, Nebel MC, et al. Continuous release of hepatitis C virus (HCV) by peripheral blood mononuclear cells and B-lymphoblastoid cell-line cultures derived from HCV-infected patients. J Gen Virol 2005; 86: 1717-1727. Links
22 Ducoulombier D, Roque-Afonso AM, Di Liberto G, Penin F, Kara R, Richard Y, et al. Frequent compartmentalization of hepatitis C virus variants in circulating B cells and monocytes. HEPATOLOGY 2004; 39: 817-825. Links
23 Sung VM, Shimodaira S, Doughty AL, Picchio GR, Can H, Yen TS, et al. Establishment of B-cell lymphoma cell lines persistently infected with hepatitis C virus in vivo and in vitro: the apoptotic effects of virus infection. J Virol 2003; 77: 2134-2146. Links
24 Negri E, Little D, Boiocchi M, La Vecchia C, Franceschi S. B-cell non-Hodgkin's lymphoma and hepatitis C virus infection: a systematic review. Int J Cancer 2004; 111: 1-8. Links
25 Zuckerman E, Zuckerman T, Levine AM, Douer D, Gutekunst K, Mizokami M, et al. Hepatitis C virus infection in patients with B-cell non-Hodgkin lymphoma. Ann Intern Med 1997; 127: 423-428. Links
26 Gisbert JP, Garcia-Buey L, Pajares JM, Moreno-Otero R. Systematic review: regression of lymphoproliferative disorders after treatment for hepatitis C infection. Aliment Pharmacol Ther 2005; 21: 653-662. Links
27 El-Serag HB, Hampel H, Yeh C, Rabeneck L. Extrahepatic manifestations of hepatitis C among United States male veterans. HEPATOLOGY 2002; 36: 1439-1445. Links
28 Poynard T, Cacoub P, Ratziu V, Myers RP, Dezailles MH, Mercadier A, et al. Fatigue in patients with chronic hepatitis C. J Viral Hepat 2002; 9: 295-303. Links
29 Radkowski M, Wilkinson J, Nowicki M, Adair D, Vargas H, Ingui C, et al. Search for hepatitis C virus negative-strand RNA sequences and analysis of viral sequences in the central nervous system: evidence of replication. J Virol 2002; 76: 600-608. Links
30 Laskus T, Radkowski M, Bednarska A, Wilkinson J, Adair D, Nowicki M, et al. Detection and analysis of hepatitis C virus sequences in cerebrospinal fluid. J Virol 2002; 76: 10064-10068. Links
31 Pol S, Driss F, Devergie A, Brechot C, Berthelot P, Gluckman E. Is hepatitis C virus involved in hepatitis-associated aplastic anemia? Ann Intern Med 1990; 113: 435-437. Links
32 Brown KE, Tisdale J, Barrett AJ, Dunbar CE, Young NS. Hepatitis-associated aplastic anemia. N Engl J Med 1997; 336: 1059-1064. Links
33 Sangar D, Carroll A. A tale of two strands: reverse-transcriptase polymerase chain reaction detection of hepatitis C virus replication. HEPATOLOGY 1998; 28: 11731176. Links
34 Komurian-Pradel F, Perret M, Deiman B, Sodoyer M, Lotteau V, Paranhos-Baccala G, et al. Strand specific quantitative real-time PCR to study replication of hepatitis C virus genome. J Virol Methods 2004; 116: 103-106. Links
35 Laskus T, Radkowski M, Jablonska J, Kibler K, Wilkinson J, Adair D, et al. Human immunodeficiency virus facilitates infection/replication of hepatitis C virus in native human macrophages. Blood 2004; 103: 3854-3859. Links
36 Castet V, Fournier C, Soulier A, Brillet R, Coste J, Larrey D, et al. Alpha interferon inhibits hepatitis C virus replication in primary human hepatocytes infected in vitro. J Virol 2002; 76: 8189-8199. Links
37 Vona G, Tuveri R, Delpuech O, Vallet A, Canioni D, Ballardini G, et al. Intrahepatic hepatitis C virus RNA quantification in microdissected hepatocytes. J Hepatol 2004; 40: 682-688. Links
38 Gosalvez J, Rodriguez-Inigo E, Ramiro-Diaz JL, Bartolome J, Tomas JF, Oliva H, Carreno V. Relative quantification and mapping of hepatitis C virus by in situ hybridization and digital image analysis. HEPATOLOGY 1998; 27: 1428-1434. Links
39 Myers TW, Gelfand DH. Reverse transcription and DNA amplification by a Thermus thermophilus DNA polymerase. Biochemistry 1991; 30: 7661-7666. Links
40 Blackard J, Smeaton L, Hiasa Y, Horiike N, Onji M, Jamieson D, et al. Detection of hepatitis C virus (HCV) in serum and peripheral blood mononuclear cells of HCV-monoinfected and HIV/HCV-coinfected persons. J Infect Dis 2005; 192: 258-265. Links
41 Hiasa Y, Blackard J, Lin W, Kamegaya Y, Horiike N, Onji M, et al. Cell-based models of sustained, interferon-sensitive hepatitis C virus genotype 1 replication. J Virol Methods 2006; 132: 195-203. Links
42 Lohmann V, Korner F, Koch J, Herian U, Theilmann L, Bartenschlager R. Replication of subgenomic hepatitis C virus RNAs in a hepatoma cell line. Science 1999; 285: 110-113. Links
43 Blight K, Kolykhalov A, Rice C. Efficient initiation of HCV RNA replication in cell culture. Science 2000; 290: 1972-1974. Links
44 Bukh J, Pietschmann T, Lohmann V, Krieger N, Faulk K, Engle R, et al. Mutations that permit efficient replication of hepatitis C virus RNA in Huh-7 cells prevent productive replication in chimpanzees. Proc Natl Acad Sci U S A 2002; 99: 14416-14421. Links
45 Heller T, Saito S, Auerbach J, Williams T, Moreen T, Jazwinski A, et al. An in vitro model of hepatitis C virion production. Proc Natl Acad Sci U S A 2005; 102: 2579-2583. Links
46 Wakita T, Pietschmann T, Kato T, Date T, Miyamoto M, Zhao Z, et al. Production of infectious hepatitis C virus in tissue culture from a cloned viral genome. Nat Med 2005; 11: 791-796. Links
47 Zhong J, Gastaminza P, Cheng G, Kapadia S, Kato T, Burton D, et al. Robust hepatitis C virus infection in vitro. Proc Natl Acad Sci U S A 2005; 102: 9294-9299. Links
48 Yi M, Villanueva RA, Thomas DL, Wakita T, Lemon SM. Production of infectious genotype 1a hepatitis C virus (Hutchinson strain) in cultured human hepatoma cells. Proc Natl Acad Sci U S A 2006. Links
49 Dash S, Halim AB, Tsuji H, Hiramatsu N, Gerber MA. Transfection of HepG2 cells with infectious hepatitis C virus genome. Am J Pathol 1997; 151: 363-373. Links
50 Hiramatsu N, Dash S, Gerber MA. HCV cDNA transfection to HepG2 cells. J Viral Hepat 1997; 4 ( Suppl 1): 61-67. Links
51 Seong YR, Lee CH, Im DS. Characterization of the structural proteins of hepatitis C virus expressed by an adenovirus recombinant. Virus Res 1998; 55: 177-185. Links
52 Kalkeri G, Khalap N, Akhter S, Garry RF, Fermin CD, Dash S. Hepatitis C viral proteins affect cell viability and membrane permeability. Exp Mol Pathol 2001; 71: 194-208. Links
53 Chung R, He W, Saquib A, Contreras A, Xavier R, Chawla A, et al. Hepatitis C virus replication is directly inhibited by INF-a in a full-length binary expression system. Proc Natl Acad Sci U S A 2001; 98: 9847-9852. Links
54 Wolk B, Gremion C, Ivashkina N, Engler OB, Grabscheid B, Bieck E, et al. Stable human lymphoblastoid cell lines constitutively expressing hepatitis C virus proteins. J Gen Virol 2005; 86: 1737-1746. Links
55 Bare P, Massud I, Belmonte L, Corti M, Villafane M, Perez Bianoco R, et al. HCV recovery from peripheral blood mononuclear cell culture supernatants derived from HCV-HIV co-infected haemophilic patients with undetectable HCV viraemia. Haemophilia 2003; 9: 598-604. Links
56 Pham TN, Macparland SA, Coffin CS, Lee SS, Bursey FR, Michalak TI. Mitogen-induced upregulation of hepatitis C virus expression in human lymphoid cells. J Gen Virol 2005; 86: 657-666. Links
57 Cribier B, Schmitt C, Bingen A, Kirn A, Keller F. In vitro infection of peripheral blood mononuclear cells by hepatitis C virus. J Gen Virol 1995; 76(Pt 10): 2485-2491. Links
58 Muller HM, Pfaff E, Goeser T, Kallinowski B, Solbach C, Theilmann L. Peripheral blood leukocytes serve as a possible extrahepatic site for hepatitis C virus replication. J Gen Virol 1993; 74(Pt 4): 669-676. Links
59 Caussin-Schwemling C, Schmitt C, Stoll-Keller F. Study of the infection of human blood derived monocyte/macrophages with hepatitis C virus in vitro. J Med Virol 2001; 65: 14-22. Links
60 Radkowski M, Bednarska A, Horban A, Stanczak J, Wilkinson J, Adair D, Nowicki M, et al. Infection of primary human macrophages with hepatitis C virus in vitro: induction of tumour necrosis factor-a and interleukin 8. J Gen Virol 2004; 85: 47-59. Links
61 Sansonno D, Iacobelli AR, Cornacchiulo V, Iodice G, Dammacco F. Detection of hepatitis C virus (HCV) proteins by immunofluorescence and HCV RNA genomic sequences by non-isotopic in situ hybridization in bone marrow and peripheral blood mononuclear cells of chronically HCV-infected patients. Clin Exp Immunol 1996; 103: 414-421. Links
62 Chang M, Marquardt AP, Wood BL, Williams O, Cotler SJ, Taylor SL, et al. In situ distribution of hepatitis C virus replicative-intermediate RNA in hepatic tissue and its correlation with liver disease. J Virol 2000; 74: 944-955. Links
63 Muratori L, Gibellini D, Lenzi M, Cataleta M, Muratori P, Morelli MC, et al. Quantification of hepatitis C virus-infected peripheral blood mononuclear cells by in situ reverse transcriptase-polymerase chain reaction. Blood 1996; 88: 2768-2774. Links
64 Revie D, Braich RS, Bayles D, Chelyapov N, Khan R, Geer C, et al. Transmission of human hepatitis C virus from patients in secondary cells for long term culture. Virol J 2005; 2: 37. Links
65 Sansonno D, Lauletta G, Dammacco F. Detection and quantitation of HCV core protein in single hepatocytes by means of laser capture microdissection and enzyme-linked immunosorbent assay. J Viral Hepat 2004; 11: 27-32. Links
66 Favre D, Muellhaupt B. Potential cellular receptors involved in hepatitis C virus entry into cells. Lipids Health Dis 2005; 4: 9-14. Links
67 Laskus T, Radkowski M, Piasek A, Nowicki M, Horban A, Cianciara J, et al. Hepatitis C virus in lymphoid cells of patients coinfected with human immunodeficiency virus type 1: evidence of active replication in monocytes/macrophages and lymphocytes. J Infect Dis 2000; 181: 442-448. Links
68 Laskus T, Radkowski M, Wang L, Vargas H, Rakela J. Search for hepatitis C virus extrahepatic replication sites in patients with acquired immunodeficiency syndrome: specific detection of negative-strand viral RNA in various tissues. HEPATOLOGY 1998; 28: 1398-1401. Links
69 Laskus T, Radkowski M, Wang L, Jang S, Vargas H, Rakela J. Hepatitis C virus quasispecies in patients infected with HIV-1: correlation with extrahepatic replication. Virology 1998; 248: 164-171. Links
70 Radkowski M, Kubicka J, Kisiel E, Cianciara J, Nowicki M, Rakela J, Laskus T. Detection of active hepatitis C virus and hepatitis G virus/GB virus C replication in bone marrow in human subjects. Blood 2000; 95: 3986-3989. Links
71 Laskus T, Radkowski M, Wang L, Vargas H, Rakela J. The presence of active hepatitis C virus replication in lymphoid tissue in patients coinfected with human immunodeficiency virus type 1. J Infect Dis 1998; 178: 1189-1192. Links
72 Nowicki MJ, Laskus T, Nikolopoulou G, Radkowski M, Wilkinson J, Du WB, et al. Presence of hepatitis C virus (HCV) RNA in the genital tracts of HCV/HIV-1-coinfected women. J Infect Dis 2005; 192: 1557-1565. Links
73 Bouffard P, Hayashi PH, Acevedo R, Levy N, Zeldis JB. Hepatitis C virus is detected in a monocyte/macrophage subpopulation of peripheral blood mononuclear cells of infected patients. J Infect Dis 1992; 166: 1276-1280. Links
74 Gabrielli A, Manzin A, Candela M, Caniglia ML, Paolucci S, Danieli MG, et al. Active hepatitis C virus infection in bone marrow and peripheral blood mononuclear cells from patients with mixed cryoglobulinaemia. Clin Exp Immunol 1994; 97: 87-93. Links
75 Gil B, Qian C, Riezu-Boj JI, Civeira MP, Prieto J. Hepatic and extrahepatic HCV RNA strands in chronic hepatitis C: different patterns of response to interferon treatment. HEPATOLOGY 1993; 18: 1050-1054. Links
76 Navas S, Castillo I, Bartolome J, Marriott E, Herrero M, Carreno V. Positive and negative hepatitis C virus RNA strands in serum, liver and peripheral blood mononuclear cells in anti-HCV patients: relation with the liver lesion. J Hepatol 1994; 21: 182-186. Links
77 Willems M, Peerlinck K, Moshage H, Deleu I, Van den Eynde C, Vermylen J. Hepatitis C virus-RNAs in plasma and in peripheral blood mononuclear cells of hemophiliacs with chronic hepatitis C: evidence for viral replication in peripheral blood mononuclear cells. J Med Virol 1994; 42: 272-278. Links
78 Radkowski M, Wang LF, Vargas HE, Rakela J, Laskus T. Detection of hepatitis C virus replication in peripheral blood mononuclear cells after orthotopic liver transplantation. Transplantation 1998; 66: 664-666. Links
79 Goutagny N, Fatmi A, De Ledinghen V, Penin F, Couzigou P, Ichauspe G. Evidence of viral replication in circulating dendritic cells during hepatitis C virus infection. J Infect Dis 2003; 187: 1951-1958. Links
80 Radkowski M, Gallegos-Orozco J, Jablonska J, Colby T, Walewska-Zielecka B, Kubicka J, et al. Persistence of hepatitis C virus in patients successfully treated for chronic hepatitis C. HEPATOLOGY 2005; 41: 106-114. Links
81 Lerat H, Rumin S, Habersetzer F, Berby F, Trabaud M, Trepo C, et al. In vivo tropism of hepatitis C virus genomic sequences in hematopoietic cells: influence of viral load, viral genotype, and cell phenotype. Blood 1998; 91: 3841-3849. Links
82 Shimizu Y, Igarashi H, Kanematu T, Fujiwara K, Wong D, Purcell R, et al. Sequence analysis of the hepatitis C virus genome recovered from serum, liver, and peripheral blood mononuclear cells of infected chimpanzees. J Virol 1997; 71: 5769-5773. Links
83 Lerat H, Shimizu Y, Lemon S. Cell type-specific enhancement of hepatitis C virus internal ribosome entry site-directed translation due to 5 nontranslated region substitutions selected during passage of virus in lymphoblastoid cells. J Virol 2000; 74: 7024-7031. Links
84 Shimizu Y, Iwamtoto A, Hijikata M, Purcell R, Yoshikura H. Evidence for in vitro replication of hepatitis C virus genome in a T-cell line. Proc Natl Acad Sci U S A 1992; 89: 5477-5481. Links
85 Nakajima N, Hijikata M, Yoshikura H, Shimizu Y. Characterization of long-term cultures of hepatitis C virus. J Virol 1996; 70: 3325-3329. Links
86 Kato N, Ikeda M, Sugiyama K, Mizutani T, Tanaka T, Shimotohno K. Hepatitis C virus population dynamics in human lymphocytes and hepatocytes infected in vitro. J Gen Virol 1998; 79(Pt 8): 1859-1869. Links
87 Roque-Afonso A, Ducoulombier D, Di Liberto G, Kara R, Gigou M, Dussaix E, et al. Compartmentalization of hepatitis C virus genotypes between plasma and peripheral blood mononuclear cells. J Virol 2005; 79: 6349-6357. Links
88 Roque-Afonso A, Jiang J, Penin F, Tareau C, Samuel D, Petit M, et al. Nonrandom distribution of hepatitis C virus quasispecies in plasma and peripheral blood mononuclear cell subsets. J Virol 1999; 73: 9213-9221. Links
89 Zehender G, De Maddalena C, Bernini F, Ebranati E, Monti G, Pioltelli P, et al. Compartmentalization of hepatitis C virus quasispecies in blood mononuclear cells of patients with mixed cryoglobulinemic syndrome. J Virol 2005; 79: 9145-9156. Links
90 Maggi F, Fornai C, Vatteroni M, Giorgi M, Morrica A, Pistello M, et al. Differences in hepatitis C virus quasispecies composition between liver, peripheral blood mononuclear cells and plasma. J Gen Virol 1997; 78: 1521-1525. Links
91 Maggi F, Fornai C, Morrica A, Vatteroni M, Giorgi M, Marchi S, Ciccorossi P, et al. Divergent evolution of hepatitis C virus in liver and peripheral blood mononuclear cells of infected patients. J Med Virol 1999; 57: 57-63. Links
92 Laskus T, Radkowski M, Wang L, Nowicki M, Rakela J. Uneven distribution of hepatitis C virus quasispecies in tissues from subjects with end-stage liver disease: confounding effect of viral adsorption and mounting evidence for the presence of low-level extrahepatic replication. J Virol 2000; 74: 1014-1017. Links
93 Okuda M, Hino K, Korenaga M, Yamaguchi Y, Katoh Y, Okita K. Differences in hypervariable region 1 quasispecies of hepatitis C virus in human serum, peripheral blood mononuclear cells, and liver. HEPATOLOGY 1999; 29: 217-222. Links
94 Navas S, Martin J, Quiroga J, Castillo I, Carreno V. Genetic diversity and tissue compartmentalization of the hepatitis C virus genome in blood mononuclear cells, liver, and serum from chronic hepatitis C patients. J Virol 1998; 72: 1640-1646. Links
95 Jang S, Wang L, Radkowski M, Rakela J, Laskus T. Differences between hepatitis C virus 5 untranslated region quasispecies in serum and liver. J Gen Virol 1999; 80: 711-716. Links
96 Forton DM, Karayiannis P, Mahmud N, Taylor-Robinson SD, Thomas HC. Identification of unique hepatitis C virus quasispecies in the central nervous system and comparative analysis of internal translational efficiency of brain, liver, and serum variants. J Virol 2004; 78: 5170-5183. Links
97 Kao J, Chen P, Lai M, Wang T, Chen D. Positive and negative strand of hepatitis C virus RNA sequences in peripheral blood mononuclear cells in patients with chronic hepatitis C: no correlation with viral genotypes 1b, 2a, and 2b. J Med Virol 1997; 52: 270-274. Links
98 Mazur W, Mazurek U, Jurzak M, Wilczok T, Bulanowski Z, Gonciarz Z. Positive and negative strands of HCV-RNA in sera and peripheral blood mononuclear cells of chronically hemodialyzed patients. Med Sci Monit 2001; 7: 108-115. Links
99 Taliani G, Badolato M, Lecce R, Poliandri G, Bozza A, Duca F, et al. Hepatitis C virus RNA in peripheral blood mononuclear cells: relation with response to interferon treatment. J Med Virol 1995; 47: 16-22. Links
100 Saleh M, Tibbs C, Koskinas J, Pereira L, Bomford A, Portmann B, et al. Hepatic and extrahepatic hepatitis C virus replication in relation to response to interferon therapy. HEPATOLOGY 1994; 20: 1399-1404. Links
101 Castillo I, Rodriguez-Inigo E, Bartolome J, de Lucas S, Ortiz-Movilla N, Lopez-Alcorocho J, Pardo M, et al. Hepatitis C virus replicates in peripheral blood mononuclear cells of patients with occult hepatitis C infection. Gut 2005; 54: 682-685. Links
102 Pham T, MacParland S, Mulrooney P, Cooksley H, Naoumov N, Michalak T. Hepatitis C virus persistence after spontaneous or treatment-induced resolution of hepatitis C. J Virol 2004; 78: 5867-5874. Links
103 Gong G, Lai L, Jiang Y, He Y, Su X. HCV replication in PBMC and its influence on interferon therapy. World J Gastroenterol 2003; 9: 291-294. Links
104 Watkins-Riedel T, Ferenci P, Steindl-Munda P, Gschwantler M, Mueller C, Woegerbauer M. Early prediction of hepatitis C virus (HCV) infection relapse in nonresponders to primary interferon therapy by means of HCV RNA whole-blood analysis. Clin Infect Dis 2004; 39: 1754-1760. Links
105 Schmidt W, Wu P, Brashear D, Klinzman D, Phillips M, LaBrecque D, Stapleton J. Effect of interferon therapy on hepatitis C virus RNA in whole blood, plasma, and peripheral blood mononuclear cells. HEPATOLOGY 1998; 28: 1110-1116. Links
106 Manavi M, Baghestanian M, Watkins-Riedel T, Battistutti W, Pischinger K, Schatten C, et al. Detection of hepatitis C virus (HCV) RNA in normal cervical smears of HCV-seropositive patients. Clin Infect Dis 2002; 35: 966-973. Links
107 Vassilopoulos D, Calabrese LH. Extrahepatic immunological complications of hepatitis C virus infection. AIDS 2005; 19 Suppl 3: S123-127. Links
108 Ray RB, Lagging LM, Meyer K, Steele R, Ray R. Transcriptional regulation of cellular and viral promoters by the hepatitis C virus core protein. Virus Res 1995; 37: 209-220. Links
109 Lanford R, Chavez D, Chisari F, Sureau C. Lack of detection of negative-strand hepatitis C virus RNA in peripheral blood mononuclear cells and other extrahepatic tissues by the highly strand-specific rTth reverse transcriptase PCR. J Virol 1995; 69: 8079-8083. Links
110 Boisvert J, He X, Cheung R, Keeffe E, Wright T, Greenberg H. Quantitative analysis of hepatitis C virus in peripheral blood and liver: replication detected only in liver. J Infect Dis 2001; 184: 827-835. Links
111 Laskus T, Radkowski M, Wang L, Cianciara J, Vargas H, Rakela J. Hepatitis C virus negative strand RNA is not detected in peripheral blood mononuclear cells and viral sequences are identical to those in serum: a case against extrahepatic replication. J Gen Virol 1997; 78: 2747-2750. Links
112 Mazurek U, Wilczok T, Mazur W, Bednarek I, Strzalka B, Jurzak M, et al. Distribution of hepatitis C virus RNA in whole blood of patients with HCV infection and HBV-HCV coinfection. Med Sci Monit 2002; 8: CR125-130. Links
113 Lerat H, Berby F, Trabaud MA, Vidalin O, Major M, Trepo C, et al. Specific detection of hepatitis C virus minus strand RNA in hematopoietic cells. J Clin Invest 1996; 97: 845-851. Links
114 Lin L, Fevery J, Yap S. A novel strand-specific RT-PCR for detection of hepatitis C virus negative-strand RNA (replicative intermediate): evidence of absence or very low level of HCV replication in peripheral blood mononuclear cells. J Virol Methods 2002; 100: 97-105. Links
115 Mellor J, Haydon G, Blair C, Livingstone W, Simmonds P. Low level or absent in vivo replication of hepatitis C virus and hepatitis G virus/GB virus C in peripheral blood mononuclear cells. J Gen Virol 1998; 79(Pt 4): 705-714. Links
|
|
| |
| |
|
|
|